エビデンス
Evidence- マグネシウム
- 健康
栄養素代謝におけるマグネシウムの重要性
Bull. Soc. Sea Water Sci., Jpn., 64, 202 - 210(2010)
Bulletin of the Society of Sea Water Science, Japan
特集「「にがり」を中心としたマグネシウムの食品栄養学的研究」 (総説)
栄養素代謝におけるマグネシウムの重要性
上原 万里子*1,石島 智子*2,松本 一朗*2,岡田 晋司*2,荒井 綜一*3,
阿部 啓子*2,勝間田 真一*1,松崎 広志*4,鈴木 和春*1
Importance of Magnesium in Nutrient Metabolism
Mariko UEHARA *1 , Tomoko ISHIJIMA *2 , Ichiro MATSUMOTO *2 , Shinji OKADA *2 , Soichi ARAI *3 , Keiko ABE *2 , Shin-ichi KASUMATA *1 , Hiroshi MATSUZAKI *4 and Kazuharu SUZUKI *1
Magnesium (Mg) is essential for a diverse range of physiological functions, as it is involved in a variety of the body’s biochemical processes.Mg defi ciency, either from inadequate intake or from excess excretion, is often suspected to be associated with the development of many symptoms and diseases. The nutritional and physiological importance of Mg has already been well established. Mg was fi nally confi rmed to be the most effective element for preventing several diseases, such as cardiovascular disease and metabolic disorders including obesity, diabetes and hypertension. Recently, many studies and meta-analyses have proved the above fi ndings. Furthermore, a number of studies have reported the effects of Mg defi ciency on carbohydrate, lipid, protein, vitamin and mineral metabolism. In this review, we discuss the effects of dietary-Mg defi ciency on proteins, lipids, ascorbic acid and mineral metabolism, including kidney calcifi cation and bone loss, in rodents. Furthermore, we performed transcriptome analysis to comprehensively understand the effects of dietary-Mg defi ciency in rat livers and femora by using DNA microarray. Key Words : Magnesium, Protein, Lipid, Ascorbic Acid, DNA microarray
1.はじめに
マグネシウム(Mg)は生体にとって必須なミネラルであり,生命維持にとって根本的な生体内反応に関与するために欠くことのできない栄養素である.とりわけ,エネルギー代謝においてはアデノシン三リン酸(ATP)生成と消費におけるリン酸基の転移反応における触媒として必要とされており(Fig.1)1),解糖系2)およびイオンポンプと神経や筋などの興奮性細胞の活動1, 3),さらには細胞内シグナル伝達においてもMg が重要な役割を果たしている4).生命維持に関わるMg が欠乏すると多様な臨床的兆候が現れることはよく知られている.罹患率とMg 摂取量について,症状の有無と血清Mg 濃度の変動など,様々な疾病とMg の関係が検討された結果,Mg による疾病発症のリスク低減効果がメタアナリシスによって立証されつつある.そのような疾患には糖尿病や高血圧などのメタボリックシンドローム,急性虚血性発作,心筋梗塞などが挙げられる5-7).Mg 欠乏と5 大栄養素の代謝に関して多くの報告がなされているが,本総説では,実験動物を用いた試験により,Mg 欠乏とたんぱく質,脂質,アスコルビン酸,他のミネラル,特にリン(P)との関連について,著者らの既報を中心に概説し,DNA マイクロアレイを用いMg 欠乏ラット肝および骨で変動する遺伝子発現の網羅的解析結果の概要を紹介する。

2.マグネシウム欠乏とたんぱく質代謝8)
Mg 欠乏により飼料摂取量の低下および体重増加量の減少が引き起こされ,ラットの成長は阻害される.この成長阻害,即ちMg 欠乏による体重減少は,たとえMg 正常食を摂取しているラットの摂食量をMg 欠乏ラットと揃えたとしても引き起こされることから,Mg 欠乏状態では,たんぱく質代謝が円滑に行われていないことが推察される.被験動物として幼若雄性ラットを,AIN-93G 組成の精製飼料をベースとしたMg 正常食(C)群とMg 欠乏食(MD)群に分け,28 日間の飼育観察を行った.Mg 欠乏食群の半数をMg 欠乏からの回復(R)群とし,Mg 正常食を7 日もしくは14 日間摂取させた.28 日後,Mg 欠乏食投与により最終体重(Fig.2),体重増加量,飼料効率は低下した.Mg 吸収量および体内保留量,血清Mg 濃度(Fig.2)も低値を示した.更に,Mg 欠乏食群では,尿中窒素(N)排泄量が増加し,N 体内保留量や血清アルブミン濃度も低下した(Fig.3).Mg 欠乏食からMg 正常食に切り換えた7
日もしくは14 日目の回復群では,Mg 吸収量および体内保留量,血清Mg 濃度は増加したが,体重および飼料効率についてはMg 欠乏食群との間に有意な差は認められなかった(Fig.2).N の体内保留量は回復14 日目で,血清アルブミン濃度では回復7 日および14 日目で,Mg 欠乏食群よりも高値を示した(Fig.3).以上の結果より,Mg欠乏状態ではたんぱく質利用が障害され,Mg 正常食投与により一部は回復されるが,成長遅延については,14日間の期間では完全に回復しないことが示唆された。
3.マグネシウム欠乏と過酸化脂質9)
Mg 欠乏は脂質代謝にも影響を及ぼし,生体膜の脂質過酸化に関連した組織傷害を引き起こすことが報告されている10-12)。
Mg 欠乏においては,過酸化脂質から生成されるマロンジアルデヒド(MDA)を意味するチオバルビツール酸反応物質(TBARS)の値の増加が観察されているがその酸化傷害の詳細については不明な点が多い。
そこで,筆者らはMg 欠乏により,生体膜の過酸化一次生成物であるphosphatidylcholine hydroperoxide (PCOOH)およびphosphatidylethanolamine hydroperoxide (PEOOH)の変動とMg,カルシウム(Ca),鉄(Fe),銅(Cu)の組織分布状態との関連を検討した。
4.マグネシウム欠乏へのアスコルビン酸投与と脂質代謝
Hus ら18)はMg 欠乏ラットにおいて,肝臓のアスコルビン酸(AsA)濃度の低下を報告している.AsA は多くの生体内反応に重要な役割を担っており,抗酸化物質としてもよく知られている.AsA はα- トコフェロールと協調して抗酸化作用を発揮する.これらのことから、Mg 欠乏による肝臓AsA 濃度の低下は,Mg 欠乏ラットの脂質過酸化にも影響を及ぼすことが考えられる.
筆者らは,Mg 欠乏ラットではAsA の要求量が増加していることを推測し,Mg 欠乏ラットへのAsA 投与の影響について検討した.被験動物として幼若雄性ラットを用い,AIN-93G をベースとしたMg 正常食の対照群,Mg 欠乏群,Mg 欠乏+AsA 投与群の3 群に分け,42 日間の飼育観察を行った.


幼若雄性ラットを用い,AIN-93G をベースとしたMg 欠乏食群と正常Mg で摂食量をMg 欠乏食群に揃えた対照群の2 群に分け,28 日間の飼育観察を行った.その結果,Mg 欠乏食投与により,肝臓中のFe 濃度は増加した.Fe はFenton もしくはHarber-Weiss 反応のような脂質過酸化の開始反応に重要であることから,この増加がMg 欠乏による脂質過酸化の一因と考えられた。
その2 つのメカニズムとして,
1)腸管でのFe 吸収はMg の存在により障害されるので,Mg 欠乏時
にはFe 吸収が上昇する,
2)Mg 欠乏では溶血が起こり,ヘモグロビンからのFe の遊離が促進されることが考えられる.多くの研究者により,肝臓におけるFe 濃度の上昇はMDA そのものやTBARSとの相関性が報告されている10- 12).
筆者らはMg 欠乏による肝臓のFe とPCOOH の上昇を観察し,両者の間には正の相関(r=0.837,p=0.001)があることを確認している(Fig.4).また心臓においてもFe とPCOOH(r=0.780,p=0.003),CaとPCOOH(r= 0.633,p=0.027)の間に,それぞれ正の相関が認められている(Fig.4).多くの動物実験,疫学調査,臨床試験により,Mg欠乏は冠動脈疾患の発症に関連のあることがわかっている。Mg 欠乏状態では,心筋肉細胞内Caの上昇が観察され,それが脂質過酸化の一因となっている。
筆者らの研究では,大動脈におけるPCOOH の増加も観察している。
この時,動脈におけるミネラル測定はしていないが,赤血球膜におけるCa,Fe,Cu濃度,血漿中のCu濃度が増加していた。
このことは大動脈の粘膜においても同様なことが起こる可能性を示唆している筆者らは以前に鉄欠乏ラット肝におけるCu 濃度とPCOO.H の増加を観察している13)。
Mg 欠乏ラット肝においてもCu 濃度とPCOOH の間には有意な正の相関(r = 0.579,p = 0.048)が認められている(Fig.4)。
フリーラジカルはCu2+ の存在によっても反応性の強い形態に変化する.Gueux ら10)は,脂質異常症はMg 欠乏ラットにおいて観察され,それは,Mg 欠乏によるVLDL およびLDL の増加によるものとしている.また,invitro 試験のCu イオンが誘導するLDL酸化は,Mg 欠乏状態において更にその感受性が高まるものとしている10).筆者らはMg 欠乏ラット赤血球膜および血漿におけるCu 濃度とPCOOH も正の相関(r=0.784,p=0.008およびr=0.907,p=0.0001)を示し 、赤血球膜ではPEOOH の増加も観察した.他の組織におけるPEOOH 濃度の変化は認められなかった。筆者らは以前にMg 欠乏ラットの腎石灰化などの腎障害および機能不全について報告している14).このことからMg 欠乏ラット腎におけるPCOOH の増加を予測したが,変動はみられなかった。Mg 欠乏ラットの腎臓ではCa およびCu 濃度が増加していたが,これらは遊離型ではなく,Ca はリン酸カルシウムなどの形態を,Cu については体内の金属補足たんぱく質であるメタロチオネインなどと結合していることにより,腎臓における脂質過酸化には寄与しなかったものと思われる.しかし,この点については推測の域を出ず,更なる検討が必要である.
Mg 欠乏における別の角度からの脂質過酸化の亢進は,抗酸化系の障害が考えられ,superoxide dismutase( SOD),catalase およびglutathione peroxidase (GPX)などの抗酸化酵素の活性低下が報告されている15, 16).筆者らの試験におけるMg 欠乏による抗酸化酵素活性は変化しなかった
が,DNA 傷害の指標である尿中8-hydroxy-deoxiguanosin(8-OH-dG)濃度はMg 欠乏ラットで対照ラットの2 倍値を示した.この結果は,Mg 欠乏ラットの体内でDNA 傷害が起こっていることを間接的に示すものであるが,組織の8-OH-dG 量についても測定し,確認する必要がある.
4.マグネシウム欠乏へのアスコルビン酸投与と脂質代謝
Hus ら18)はMg 欠乏ラットにおいて,肝臓のアスコル
ビン酸(AsA)濃度の低下を報告している.AsA は多くの
生体内反応に重要な役割を担っており,抗酸化物質として
もよく知られている.AsA はα- トコフェロールと協調し
て抗酸化作用を発揮する.これらのことから,Mg 欠乏に
よる肝臓AsA 濃度の低下は,Mg 欠__乏ラットの脂質過酸化
にも影響を及ぼすことが考えられる.
筆者らは,Mg 欠乏ラットではAsA の要求量が増加して
いることを推測し,Mg 欠乏ラットへのAsA 投与の影響に
ついて検討した.被験動物として幼若雄性ラットを用い,
AIN-93G をベースとしたMg 正常食の対照群,Mg 欠乏群,
Mg 欠乏+AsA 投与群の3 群に分け,42 日間の飼育観察を
行った.

Fig.4 Correlation coeffi cients of phosphatidylcholine hydroperoxide
(PCOOH) and minerals in four tissues
その結果,Mg 欠乏状態では,他の研究結果と同様,最終体重が低下した。AsA投与群では体重,Mg 吸収および体内保留量,血清Mg 濃度に影響を与えなかった。Mg 欠乏ラットではAsA合成に関連したグルクルノラクトンまたはグロノラクトンの利用能が低下することが知られている18)。
筆者らの試験では,Mg 欠乏ラットの血清AsA 濃度の減少を観察している.これは,Mg 欠乏がAsA 合成を阻害し,体内でのAsA 要求量が増加していることを示唆する結果である.筆者らは前項において,Mg 欠乏ラット血漿でのPCOOH 量の増加を確認している.今回の42 日間飼育の場合においても,Mg 欠乏による血清PCOOH の増加が観察された.グルタチオンはAsA の酸化型から還元型への変換を触媒する19).Mg 欠乏はグルタチオン代謝に影響を及ぼすことが推察されるが,今後の検討が必要とされる。
Mg 欠乏状態へのAsA 投与により,血清AsA 濃度は正常範囲となり,Mg 欠乏によるPCOOH 増加も抑制された。
Mg 欠乏状態では,VLDL コレステロールの増加とHDLコレステロールの低下が観察されているが,これは,コレステロールをエステル化するlecithin-cholesterol acyltransferase(LCAT)の活性化にMg が必要とされることに関連している20).また、Mg 欠乏では、リポたんぱく質中のtriacylglycerol (TG)が増加し,アポたんぱく質部分が減少する21).更にAsA はコレステロール代謝においても重要な役割を担っている.AsA 欠乏モルモットを用いた試験では,血清および肝臓中のコレステロール濃度が増加するとした報告がある21).これは,AsA がコレステロールの7α-hydroxylation に関与しており,AsA 欠乏モルモットでは,コレステロールの異化が進まないためである.Mg 欠乏動物では2 次的にAsA 欠乏と同様な状態となり,コレステロール代謝が障害されることが考えられる.筆者らの試験において,Mg 欠乏ラットの血清総コレステロール(TC)とTG は増加し,AsA 投与では変化がみられなかったが,同じく肝臓でMg 欠乏により増加したコレステロールとTG は,AsA 投与により正常レベルに戻った.多くの研究報告がAsA のコレステロール低下作用を報告しているが,その詳細については検討されるべきである.既報によれば,高コレステロール血症のヒト介入試験において,一日4 g のAsA を2 ヶ月間摂取したが,血漿TC およびTG は変動しなかった22).この肝臓と血清濃度の結果の違いはAsA の投与量,投与方法,被験動物および被験者の週齢もしくは年齢の違いによるものかもしれない.
5.マグネシウム欠乏と腎石灰化に関わる因子23)
Mg 欠乏状態では,他のミネラル代謝対しても影響を及ぼす。Mg 欠乏ラットにおける腎臓の石灰化は,その1 つの典型的な例である.過去の研究によると,Mg 欠乏ラットの腎臓では,Ca とP の濃度が増加するが,筆者らの試験においては,増加した場合14)と変動がみられない場合24)とに分かれた.そこで,この差異は飼料中のP 濃度によるものと考え,現在,幼若ラットの適切な標準飼料
Table 1 Changes of serum AsA, PCOOH, TG and TC and liver TG and TC in control rats (C), Mg-defi cient rats (D) and Mg-defi cientrats supplemented with AsA (DA).
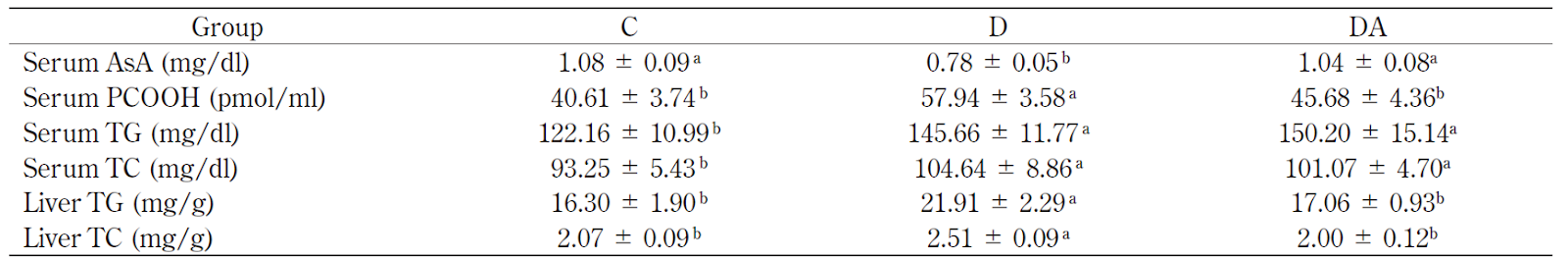
Table 2 Kidney mineral concentration and degree of nephrocalcinosis in rats fed on six experimental diets.
Normal-Mg diet Mg-defi cient diet

Values are means ± SE (n = 6). 1 Mg: effects at different dietary Mg concentrations, P: effect at different dietary P concentrations, Mg
x P : effect of interaction at p < 0.05; a,b,c Values with different superscript letters are signifi cantly different at p < 0.05; 2Score for nephrocalcinosis:
-(not detected) < + < ++ < +++ (severe).
であるAIN-93G のP 濃度を3 段階[3 (3-P),5 (5-P) or 7(7-P) g per kg diet]に変化させ,投与した.その結果,腎臓の組織学的検討において,5-P および7-P 群では腎石灰化は観察されたが,3-P では認められないことを確認した(Table 2)。またその石灰化は7-P 群で5-P 群より顕著であった(Table 2)。3-P はAIN-93G 組成の標準P 量を投与した群であり,腎石灰化は起こらなかった。5-P はAIN-76(AIN-93G より以前の標準飼料)のP 濃度を投与した群であり,たとえ,AIN-93G ベースであっても飼料中P 濃度を増加させることにより,腎石灰化が重篤となるこの結果の意味するところは,P が,腎石灰化の重要な因子と成り得るということである。この試験により,Mg 欠乏による腎石灰化は,飼料中P 濃度により発症の有無が大きく影響されることが示された。
6.マグネシウム欠乏ラット肝および大腿骨に
おけるDNA マイクロアレイ解析Mg 欠乏は,前項で述べてきたように生体内の栄養素代謝に重要な変化を引き起こす.しかし,これらのMg 欠乏で引き起される生理化学的な現象とそれらを調節する遺伝子発現変動との関係を中心とした包括的な理解には未だ至っていない。そこで,Mg 欠乏状態での生体機能に関わる様々な遺伝子発現の変動を,DNA マイクロアレイを用いて網羅的に解析することより,その代謝調節機構の全体像を把握することを試みた。
被験動物として幼若雄性ラットを用い,AIN-93G 飼料組成より酸化Mg を除いたMg 欠乏食を投与し,28 日間の飼育観察を行った.対照として正常Mg 投与群を設け,摂食量の低いMg 欠乏食投与群の摂食量に合わせた制限給餌を行った.両群より体重の揃った各3 個体の肝臓からRNA を抽出後,精製し,cDNA,cRNA を合成し,プローブ数31099 のGeneChip Rat Geneme Array 230 2.0 にハイブリダイズさせた.その後,染色およびスキャンにングを行い,Genechip Operating Software (GCOS)を用いて単解析(蛍光シグナルの数値化)と散布図の作成およびPearson の相関係数を求め,クラスター解析を行った(Fig.5).次に同様な被験動物にMg 欠乏食を投与し,35 日間の飼育観察を行った(MD35 群).対照として正常Mg 投与(C35)群を,さらに28 日間のMg 欠乏食投与後,7 日間のMg 正常食投与を行った回復(R)群も設けた.解析は同様に行った.更に,同様な幼若ラットを用い,Mg 欠乏食で6 週間飼育観察後,大腿骨を摘出,RNA 抽出後,同様にDNA マイクロアレイ解析に供した.各群間の有意差検定には正規化した31042 プローブから両群において各群3 サンプルのうち2 サンプル以上で発現していないとみなされた13729 プローブを除き,倍数変化が2 以上である1144 プローブを用いたGeneSpring GX7.3(Agilent Technologies)を用いて,シグナルを対数変換後,Student t-test を行い,False Discovery Rate (FDR) 0.05 未満を有意とした.回復過程での変化をみるためのC35 群とMD35 群およびR 群間の有意差検定には28 日間および35 日間のMg 欠乏食投与により共通で有意な変化を示したプローブを用いた.一元配置分散分析後,有意差のあるプローブについては各群間においてTukey の多重比較検定を行った。
いずれの検定においてもFDR < 0.05 を有意とした.Mg 欠乏食から正常食への飼料交換により引き起こされる各変化を示す遺伝子の機能の同定にはgene ontology(GO) stat を用いて,各変化において28 日間および35 日間のMg 欠乏食投与により共通で有意な変化を示した遺伝子群に含まれる各機能の遺伝子数に対し,その変化を示した遺伝子の多く含まれる機能の検定をFisher の正確確率検定により行い,P < 0.05 を有意とした。GeneSpring にて統計解析を行った結果,Mg 欠乏ラットの肝臓では多くの遺伝子発現が変化することが示された。また倍数変化が2 倍以上の有意な変化を示す遺伝子は31,099 のうち734 プローブと,Mg 欠乏食投与によって発現が大きく変化する遺伝子も多く,Mg 欠乏が肝臓の遺伝子発現に大きく影響していることが示唆された.Mg 欠乏により発現変化を示した遺伝子には,糖質代謝,脂質代謝,たんぱく質/アミノ酸代謝,ビタミン代謝,核酸代謝,生体異物代謝,免疫応答,輸送,細胞増殖および転写調節因子などの様々な機能に関与するものが含まれており,特に栄養素の代謝については,生理および生化学的指標の変化と結び付けられるような遺伝子の発現変化,また生化学データでは捉えることが出来ない変化など,Mg 欠乏による栄養素代謝への影響を理解する上で重要な知見が得られた(Table 3)。
三大栄養素の糖質代謝においてはMg 欠乏によりペントースリン酸回路に関連した酵素の遺伝子発現が上昇し,グリコーゲン合成酵素の遺伝子発現は低下した。脂質代謝おいては,脂肪酸合成酵素,長鎖脂肪酸の輸送,脂肪酸合成に関連する酵素の遺伝子発現は増加し,ミトコンドリアやペルオキシゾームにおけるβ酸化,コレステロールの異化に関与する酵素の遺伝子発現は低下した。
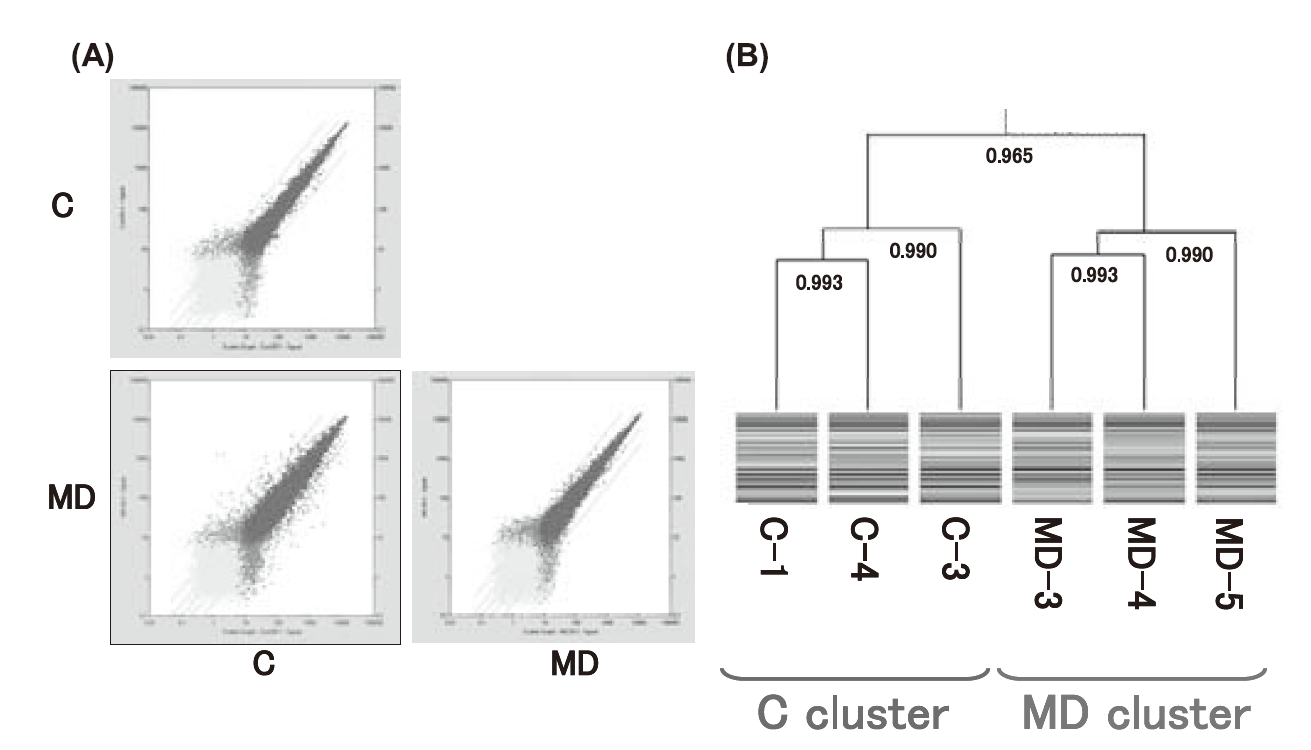
Fig.5 Scatter plotting (A), and Peason’s correlation coeffi cient analysis and cluster analysis (B)Gene expression differences were observed to a greater or lesser extent between the control (C) and the Mg-defi cient (MD) group.

また,Mg 欠乏によりSREBP-1 (sterol regulatory elementbindingprotein-1)遺伝子発現は上昇,PPARα(peroxisome proliferative activated receptor α)遺伝子発現は低下し,その標的遺伝子にも影響を及ぼした。これら転写因子の変動については,これまでに報告されておらず,新規性が高い結果である.たんぱく質代謝に関しては,ユビキチン- プロテオソーム系のプロテオリシス(たんぱく質異化)関連の遺伝子発現は上昇し,たんぱく質の翻訳開始調節に関連した遺伝子発現は低下した.アミノ酸代謝においては,システイン合成,グリシン,シスチン,システインの分解に関連した酵素の発現上昇が認められた.これらのことから体内のタンパク質量は低下することが示唆され,この体内たんぱく質量の低下が前述の体重増加量の低下すなわち成長抑制を引き起こす一因として推測された.
以上より,Mg 欠乏ラットにおける遺伝子発現の変動は栄養素代謝全般に変化をもたらし,一部の代謝に関してMg は転写調節因子の変動にも関与することが示され,Mg 欠乏により引き起こされた既知の生理生化学的現象の変化について説明可能ないくつかの知見が得られた.
次に,28 日間と35 日間のMg 欠乏食投与による影響の比較を行った.正常食投与およびMg 欠乏食投与において示される肝臓での遺伝子発現様式に28 日間と35 日間の飼育期間の違いによる差はみられず,このことからMg 欠乏食投与によって引き起こされる肝臓での遺伝子発現の変化は,28,35 日間といった長期のMg 欠乏食投与時では,ある一定の状態に保たれていることが推察された.長期間のMg 欠乏食投与時の生体内変動が,ほぼ一定の状態に保たれていることについては,たんぱく質栄養状態の生化学的解析を行った結果からも推測できる8).28 日間のMg 欠乏食投与により2 倍以上に有意な変化を示した遺伝子として734 プローブが,35 日間のMg 欠乏食投与により2 倍以上に有意な変化を示した遺伝子として637 プローブが抽出され,これらの関係をみたところ28 日間および35 日間のMg 欠乏食投与により共通で有意な発現変化示した遺伝子は357 プローブとそれぞれの飼育期間において有意差のあった遺伝子の約50 %となった(Fig.6).このうち2 倍以上の有意な変化を示す遺伝子においては,約700 または600 のプローブと,ある特定の遺伝子群を対象に一つ一つの遺伝子を比較していることから,遺伝子発現様式における28 日間および35 日間の比較とは遺伝子の数およびその条件が異なり,経時差の有無についての結果が異なったもの思われる.
35 日間のMg 欠乏食投与により発現が有意に変化した遺伝子を機能分類したところ,Mg 欠乏食投与によって影響を受けている遺伝子の機能に,28 日間と35 日間の飼育期間の違いによる差はみられなかった.また28 日間のみ,28 日間および35 日間共通,35 日間のみMg 欠乏食投与によって有意な変化を示す遺伝子を機能分類した結果,それぞれに各機能をもつ遺伝子が含まれており,遺伝子の機能に特徴はみられなかった.しかし,28 日間および35 日間共通,または35 日間のみMg 欠乏食投与によって発現変化を示した遺伝子には脂質代謝に関係する遺伝子が多く含まれていた.28 日間または35 日間のMg 欠乏食投与により発現が有意に変化した遺伝子に,脂質代謝に関係する遺伝子は多く,さらに35 日間のMg 欠乏食投与によってのみ有意な発現変化を示す遺伝子においても脂質代謝に関係する遺伝子が多いことから,35 日間のMg 欠乏食投与により発現変動を示した遺伝子の多さが,脂質代謝への影響の強さに反映していると推察された.
Table 3 Signifi cant classifi cation of up-and down regulated genes by dietary magnesium-defi ciency Metabolism Up Down Total Functions Up Down Total
Proteins 28 23 51 Transcription 16 27 43
Carbohydrates 5 10 15 Immune Response 18 * 9 27
Lipids 19** 18 37** Nerve system 10 11 21
Amino Acids 3 6 9 Circadian Rhythm 1 5 6
Vitamins 3 2 5 Transport 25 31 56
Cofactors 2 7 9 Cell Proliferation 11 8 19
DNA 3 3 6 Cell Death 13 5 18
Nucleotide 3 2 5 Cell Cycle 12 5 17
Active Oxygens 2 3 5 Cell Growth 5 4 9
Xenobiotic detoxication 1 4 5 Cell Division 6 * 2 8
Cell Homeostasis 4 3 7
* p < 0.05, ** p < 0.01 in Fisher’s exact test

Fig.6 Changed gene expressions by feeding Mg-defi cient diet
for 28 or 35 days

Fig.7 Pattern of gene expression and changed probe numbers in the recovery (R) group
DNA マイクロアレイを用いた網羅的遺伝子発現解析によりMg 欠乏からの回復過程における遺伝子発現変化を観察したところ,28 日間のMg 欠乏食投与後正常食を7 間投与したラットにおいて示される遺伝子発現様式は35日間のMg 欠乏食投与よりも正常食投与の状態に類似していることが示され,Mg 欠乏ラットにMg を適正量含有する正常食を投与し回復させることにより遺伝子発現様式は変化し,正常食投与時の発現様式に近づくことが示された.
そこで28 日間のMg 欠乏食投与後,正常食を7 日間投与したラットの肝臓において引き起こされる遺伝子の発現変化について,28 日間と35 日間のMg 欠乏食投与により共通で有意な発現変化を示す357 プローブを対象に解析を行った.28 日間のMg 欠乏食投与後,7 日間の正常食投与を行ったラットの肝臓においては,Mg 欠乏食投与によって有意な変化を示す遺伝子の約80 %が正常食投与時の遺伝子発現量へ戻る変化,つまり正常食投与時の発現への回帰を示し,そのうち約80 %の遺伝子が正常食投与時と同
程度に発現量を回復した(Fig.7).このことから,食餌性Mg 量の変動に伴い遺伝子の発現は変化することが確認された.生化学的解析においても,Mg 欠乏からの回復過程におけるミネラル利用の変化については報告18, 25)されている.また,前項で述べたようにMg 欠乏ラットにおいて引きこされるミネラル利用およびたんぱく質栄養状態の低下はMg を適正量含有する正常食の投与により改善されることが明らかになっている8).このことからMg 欠乏からの回復過程の生化学的指標の解析によって示される改善効果は,遺伝子発現の変化によって引き起こされることが推察された.
Mg 欠乏食から正常食へ飼料交換後,正常食投与時の発現量への回帰を示した遺伝子の中には,正常食投与と同程度まで未だ発現量が回復していないものや,正常食投与と同程度以上に発現が変化した遺伝子もみられた(Fig.7).またMg 欠乏食から正常食へ飼料交換後発現変化を示さなかった遺伝子もみられたことから,食餌性Mg 量の変動に伴う遺伝子の発現変化は,遺伝子によって異なることが示唆された.
Mg 欠乏食から正常食へ飼料交換後,各変動を示す遺伝子において機能分類を行った(Fig.8).その結果,Mg 欠乏食投与によって発現上昇を示した遺伝子で正常食への飼料交換後,発現変化を示さなかった遺伝子には免疫応答に関係する遺伝子が多く含まれ,食餌性Mg 量の変化に対する発現変化が遅延して引き起こされる遺伝子にも免疫応答に関係する遺伝子が多いことが示唆された.また,Mg 欠乏食投与によって発現上昇を示し,正常食への飼料交換後正常食投与時の遺伝子発現量への回帰を示した遺伝子で正常食投与と同程度以上に発現が変化した遺伝子には細胞分裂に関係する遺伝子が多く含まれ,食餌性Mg 量の変化に対する応答が鋭敏であるものには細胞分裂に関係する遺伝子が多いことが示された.また,Mg 欠乏食から正常食へ飼料交換後,正常食投与時の発現量への回帰を示した遺伝子の約80 %は機能に関係なく全体的に正常食投与時と同程度に回復していることが確認された.以上より,食餌性Mg 量の変動に伴い,肝臓で様々な栄養素代謝に関連した遺伝子発現は変化することが明らかとなった.Mg 欠乏ラットでは,骨密度および骨強度の低下,骨代謝マーカーの変動が報告されている26-28).Mg 欠乏ラット大腿骨における遺伝子発現の網羅的解析を行った結果,肝臓に比較すると変動した遺伝子の数が少なく,倍数変化も小さかったことから,1.5 倍以上で統計学的に有意な発現変動を示した遺伝子について検討した.

Fig.8 Gene expression classifi ed by biological functions in the recovery (R) group
* Signifi cant changes from Fisher exact test (p < 0.05)
まず,3.7 倍の比較的大きな上昇を示したのは,Ca sensing receptor( CaSR)遺伝子であるCasr であった.Mg はCaSR のアゴニストといわれるが,Mg 欠乏では細胞内Ca2+ 濃度が増加することも知られており,Mg欠乏によるCasr 発現の上昇により,細胞内Ca2+ 濃度増加を惹起している可能性がある29, 30).
そのため,Ca2+ が破骨細胞分化のマスター転写因子であるNFATc1 (nuclear factor activated T cells c1)の誘導および活性化を促進していることが考えられる.また,Tnfrsf6(Tumor necrosis factor receptor superfamily member 6)はMg 欠乏により1.8 倍の発現上昇を示した.Mg 欠乏ではTNF-αが上昇し,骨吸収を亢進することが知られている
31).従ってTNF の受容体ファミリーであるTnfrsf6 の発現上昇は,Mg 欠乏による骨吸収促進の一因と推察される.また,BMP (bone morphological protein)は骨芽細胞の分化と骨形成を増強する骨修飾たんぱく質であるが,Mg 欠乏により,BMP-2 inducible kinase の遺伝子であるBmp2kと BMP-3 familyの遺伝子である Gdf10( growth differentiationfactor 10)発現は約1.5 分の1 に低下した.しかし,Bmper-predicted (BMP-binding endothelial regulator-predicted)発現は4.3 倍に上昇していた.また,タンパク質リン酸化酵素(PKC)Family の遺伝子であるpkcb(β)は約2 分の1 に低下,Cox-VIC-1 発現は約2 倍に上昇,calcium/calmodulin dependent protein kinase (CaMK)の遺伝子であるCamk2b は1.5 倍に上昇,Camk2d は1.6 分の1にに低下していた.その他,骨代謝調節に重要なRANKL(receptor activator NFκB ligand),OPG(osteoprotegerin),TGF(transforming growth factor),IL(interleukin)およびIGF(insulin like growth factor)family などの遺伝子発現変動は認められなかった.
以上より,Mg 欠乏による大腿骨の遺伝子変動は,肝臓に比較すると顕著ではなかったが,一部,Ca 感知受容体,骨形成および骨吸収に関わる遺伝子発現の変動が観察された.
7.おわりに
Mg が生命維持の根本に関与し,栄養素代謝にも極めて重要な必須ミネラルであることは,様々な生理化学的研究により証明されており,1-5 項ではその一端を概説した.そして6 項において,それら代謝に関連した酵素,更には転写因子の遺伝子発現変動の網羅的解析により,Mg 欠乏により引き起こされる生理化学的な現象面を説明し得る様々な知見が得られた.Mg 摂取による生活習慣病リスク低減効果に十分な科学的根拠が示されてきた昨今,DNAマイクロアレイによるこれらの知見は,更なる科学的根拠の裏付けを示すものである.
謝 辞
本総説の第6 項は,財団法人ソルト・サイエンス研究財団の助成を受けて行った研究成果の一部である.この研究助成に対し,巻末ながら謝意を表する.
References
1) R. K. Murray, D. K. Granner, P. A. Mayes and V. W. Rodwell
(Y. Kaziro ed. and tr.),“ Harper’s Biochemistry, 25th ed.”,
pp.136-137, Maruzen Co., Ltd., Tokyo( 2003)
2) J. G. Salway( Y. Aso tr.),“ Metabolism at a Glance, 2nd ed.”,
p.5, Medical Science International Co., Ltd., Tokyo
3) R. K. Murray, D. K. Granner, P. A. Mayes and V. W. Rodwell
(Y.Kaziro ed. and tr.)“ Harper’s Biochemistry, 25th ed.”,
p.567, Maruzen Co., Ltd., Tokyo( 2003)
4) J. Takaya, H. Higashino and Y. Kobayashi, “Can magnesium
act as a second messenger? Current data on translocation
induced by various biologically active substances”, Magnes.
Res., 13( 2), pp. 139-146( 2000)
5) S. C. Larsson and A. Wolk, “Magnesium intake and risk of
type 2 diabetes: a meta-analysis”, J. Intern. Med., 262( 2), pp.
208-214( 2007)
6) S. H. Jee, E. R. 3rd Miller, E. Guallar, V. K. Singh, L. J. Appel
and M. J. Klag “The effect of magnesium supplementation
on blood pressure: a meta-analysis of randomized clinical trials”
Am. J. Hypertens. 15( 8), pp. 691-696( 2002)
7) S. Chakraborti, T. Chakraborti, M. Mandal, A. Mandal, S.
Das and S. Ghosh, “Protective role of magnesium in cardiovascular
diseases: a review”, Mol. Cel Biochem. 238( 1-2),
pp. 163-79( 2002)
8) T. Nemoto, H. Matsuzaki, M. Uehara and K. Suzuki, “Magnesium-
defi cient diet-induced reduction in protein utilization
in rats is reversed by dietary magnesium supplementation”,
Magnes. Res., 19( 1), pp. 19-27( 2006)
9) M. Uehara, H. Chiba, A. Fujii, H Matsuzaki, R Masuyama
and K. Suzuki“, Advances in Magnesium Research: Nutrition
and Health”, in Rayssiguier Y( Ed.),“ Induction of phosphatidylcholine
hydroperoxides in relation to change of tissue
mineral distribution caused by magnesium defi ciency”, pp.
291-296, John Libbey & Company Ltd, France( 2000)
10) E. Gueux, C. Cubizolles, L. Bussière, A Mazur and Y Rayssiguier,“
Oxidative modification of triglyceride-rich lipoproteins
in hypertriglyceridemic rats following magnesium
defi ciency”, Lipids, 28( 6), pp. 573-575( 1993)
11) E. Rock, C. Astier, C. Lab, X. Vignon, E Gueux, C. Motta
and Y. Rayssiguier, “Dietary magnesium deficiency in rats
enhances free radical production in skeletal muscle” J. Nutr.,
125( 5), pp. 1205-10( 1995)
12) Y. Rayssiguier, E. Gueux, L. Bussière, J. Durlach and A.
Mazur,“ Dietary magnesium affects susceptibility of lipoproteins
and tissues to peroxidation in rats”, J. Am. Coll. Nutr.,
12( 2), pp. 133-137( 1993)
13) M. Uehara, H. Chiba, H. Mogi, K. Suzuki and S. Goto, “Induction
of increased phosphatidylcholine hydroperoxide by
an iron-deficient diet in rats”, J. Nutr. Biochem., 8( 7), pp.
385-391( 1997)
14) T. Kikuchi, H. Matsuzaki, S. Sato, Y. Kajita, H. Chiba, H.
Tsuchiya, R. Masuyama, M. Uehara, K. Suzuki and S. Goto,
“Diminished kidney function and nephrocalcinosis in rats
fed a magnesium-defi cient diet”, J. Nutr. Sci. Vitaminol., 44
(4), pp. 515-23( 1998)
15) Z. Zhu, M. Kimura and Y. Itokawa, “Selenium concentration
and glutathione peroxidase activity in selenium and magnesium
defi cient rats”, Biol. Trace. Elem. Res., 37( 2-3), pp.
209-217( 1993)
16) B. P. Kumar and K. Shivakumar, “Depressed antioxidant
defense in rat heart in experimental magnesium defi ciency.
Implications for the pathogenesis of myocardial lesions”,
Biol. Trace Elem. Res., 60( 1-2), pp.139-44( 1997).
17) S. Akiyama, M. Uehara, S. Katsumata, H. Ihara, N. Hashizume
and K. Suzuki,“ Effects of dietary ascorbic acid supplementation
on lipid peroxidation and the lipid content in the
liver and serum of magnesium-defi cient rats”, Magnes. Res.,
21( 4), pp. 232-236( 2008).
18) J. M. Hsu, J. C. Jr. Smith, A. A. Yunice and G. Kepford,
“Impairment of ascorbic acid synthesis in liver extracts of
magnesium-defi cient rats”, J. Nutr., 113( 10), pp. 2041-2047
(1983)
19) J. M. Hsu, B. Rubenstein, and A. G. Paleker,“ Role of magnesium
in glutathione metabolism of rat erythrocytes”, J. Nutr.,
112( 3), pp. 488-496( 1982)
20) E. Gueux, Y. Rayssiguier, M. C. Piot and L. Alcindor,“ Reduction
of plasma lecithin-cholesterol acyltransferase activity by
acute magnesium defi ciency in the rat”, J. Nutr., 114( 8), pp.
1479-1483( 1984)
21) E. Gueux, A. Mazur, P. Cardot and Y. Rayssiguier, “Magnesium
deficiency affects plasma lipoprotein composition in
rats”, J. Nutr., 121( 8), pp. 1222-1227( 1991)
22) V. E. Peterson, P. A. Crapo, J. Weininger, H. Ginsberg and J.
Olefsky,“ Quantifi cation of plasma cholesterol and triglyceride
lavels in hypercholesterolemic subjects receiving ascorbic
acid supplements”, Am. J. Clin. Nutr., 28( 6), pp. 584-587
(1975)
22) B. Sokoloff, M. Hori, C. Saelhof, B. Mcconnell and T. Imai,
“Effect of ascorbic acid on certain blood fat metabolism factors
in animals and man”, J. Nutr., 91( 1), pp. 107-118( 1967)
23) H. Matsuzaki, S. Katsumata, M. Uehara, M. Miwa and K.
Suzuki,“ Onset of nephrocalcinosis depends on dietary phosphorus
concentration in male rats fed a magnesium-defi cient
diet”, Magnes. Res., 19( 4), pp. 255-260( 2006)
24) H. Matsuzaki, T. Nemoto M. Fuchigami, M. Uehara, K. Suzuki
and K. Nakamura, “Effects of gender on kidney function
in magnesium-defi cient rats” Biofactors, 22( 1-4): pp. 43-46
(2004)
25) Y. Rayssiguier, E. Gueux, D. Weiser,“ Effect of magnesium
deficiency on lipid metabolism in rats fed a high carbohydrate
diet”, J. Nutr., 111( 11), pp. 1876-1883( 1981)
26) R. K. Rude, F. R. Singer and H. E. Gruber,“ Skeletal and hormonal
effects of magnesium defi ciency”, J. Am. Coll. Nutr.,
28( 2), pp. 131-141( 2009).
27) M. A. Kenney, H. McCoy and L. Williams, “Effects of magnesium
defi ciency on strength, mass, and composition of rat
femur”, Calcif. Tissue Int., 54( 1), pp. 44-49( 1994)
28) S. I. Katsumata, H. Matsuzaki, R. Tsuboi, M. Uehara and K.
Suzuki, “Moderate magnesium-restricted diet affects bone
formation and bone resorption in rats”, Magnes. Res., 19( 1),
pp. 12-18( 2006)
29) E. M. Brown, G. Gamba, D. Riccardi, M. Lombardi, R. utters,
O. Kifor, A. Sun, M. A. Hediger, J. Lytton and S. C. Hebert,
“Cloning and characterization of an extracellular Ca2+-sensing
receptor from bovine parathyroid”, Nature, 366( 6455),
pp. 575-580( 1993)
30) T. Kawata and N. Nagano, “The calcium receptor and magnesium
metabolism”, Clin. Calcium, 15(11), pp. 43-50(2005)
31) R. K. Rude, H. E. Gruber, H. J. Norton, L. Y. Wei, A. Frausto
and B. G. Mills, “Bone loss induced by dietary magnesium
reduction to 10 % of the nutrient requirement in rats is associated
with increased release of substance P and tumor
necrosis factor- α”, J. Nutr., 134( 1), pp. 79-85( 2004)
2010 年7 月2 日受付
Received July 2, 2010